









Services on Demand
Article
 Article in pdf format
Article in pdf format Article in xml format
Article in xml format
 Send this article by e-mail
Send this article by e-mailBookmark
TropIKA.net
On-line version ISSN 2078-8606
TropIKA.net vol.1 no.1 Jan./Mar. 2010
Insecticide resistance in dengue vectors
Hilary RansonI,*; Joseph BurhaniII; Nongkran LumjuanII; William C Black IVIII
ILiverpool School of Tropical Medicine, UK
IIResearch Institute for Health Sciences, Chiang Mai University, Thailand
IIIColorado State University, USA
Date of database searches: October 2008.
ABSTRACT
BACKGROUND: Most national dengue control programmes rely extensively on insecticides to control the mosquito vectors of this disease.
OBJECTIVES: The objective of this review is to describe current knowledge of the extent of insecticide resistance in dengue vectors and the potential impact of this resistance on control activities.
METHODS: We searched Web of Science and PubMed for studies that included data on resistance to the four major classes of insecticides: organochlorines, carbamates, organophosphates and pyrethroids, in the dengue vectors Aedes aegypti and Aedes albopictus. Insecticide bioassay data were extracted from the published literature and the methods used to obtain, analyse and interpret this data were critically evaluated. Emphasis was placed on the two insecticide classes most widely used in dengue control, organophosphates and pyrethroids. The use of biochemical and molecular tools for resistance monitoring was also reviewed.
RESULTS: 103 studies met our inclusion criteria, of which 65 contained bioassay data which we uploaded on to a public database (IRBase). There is a strong geographical bias in published studies with nearly half originating from three countries (Thailand, India and Brazil). Bioassay data demonstrate that resistance to the organophosphate temephos and to pyrethroids is widespread in Ae. aegypti and resistance has also been reported in Ae. albopictus. Assessing the impact of insecticide resistance on vector control is complicated by variations in the methodology used to measure and report resistance, and by the lack of studies into the epidemiological consequences of insecticide resistance.
CONCLUSIONS: The lack of publicly accessible standardized data sets documenting levels of insecticide resistance in many dengue endemic countries, and the absence of studies on the operational impact of resistance, precludes a comprehensive analysis of the current global threat that insecticide resistance poses to dengue control. However, several countries with active resistance monitoring programmes have shown that insecticide resistance is reducing our ability to control dengue vectors. This situation is likely to worsen unless effective strategies are rapidly implemented to mitigate these effects.
Keywords: insecticide resistance, dengue, Aedes aegypti, Aedes albopictus, pyrethroids, temephos
Introduction
Dengue fever is a mosquito-borne febrile disease caused by any of the four dengue serotype viruses. The number of cases of dengue, dengue shock syndrome and, the most severe form, dengue haemorrhagic fever, have increased dramatically in recent years. Over 2.5 billion people live in areas where dengue viruses can be transmitted (1). Dengue is transmitted by various Aedes sp. in tropical and subtropical regions throughout the world. Aedes aegypti (L.) is the major vector but other species can be responsible for local transmission, the most important of which is Aedes albopictus. Aedes aegypti is very closely associated with the human habitat. Adult mosquitoes preferentially rest inside buildings and lay their eggs in water found around the home including water storage containers, discarded bottles and flower or plant pots. Aedes aegypti is found throughout the tropics and subtropics and its geographical range is increasing, in part due to rapid urbanization and increased global movement of people and cargo (2). Aedes albopictus originated in Asia but has been introduced into Europe, Africa and the Americas in the last 40 years. It is usually considered a less efficient dengue vector but is associated with transmission of the disease in rural and suburban areas of Southeast Asia (2).
There are no vaccines or antivirals for dengue infection and hence mosquito control is the only available means for prevention and control of the disease (3). Insecticides play an important role in dengue control. The four major classes of insecticides are organochlorines, carbamates, organophosphates and pyrethroids. Removal of mosquito breeding sites can also be very effective in areas where a reliable piped water supply is available. However, breeding sites can be small, dispersed and difficult to find. Furthermore many dengue endemic areas lack piped water making households dependent on water storage containers. These make ideal breeding sites for Aedes mosquitoes. Insecticides such as the organophosphate temephos can be applied to these large containers to reduce larval density. Temephos has very low mammalian toxicity, low odor, is available in long lasting formulations, is relatively cheap, and is usually the insecticide of choice for larviciding water supplies used for washing clothes, bathing, and cleaning dishes. It can also be used to treat drinking water although acceptance levels are often low. The insect growth regulators pryriproxyfen, novaluron and methoprene and the bacterial toxin, Bti, are also approved for use in drinking-water (4,5).
Control of adult dengue vectors by thermal fogging (spraying small particles less than 50 micrometres) or ultra-low volume sprays is only recommended in response to dengue outbreaks and even then the merits of this control method have been questioned, especially when applied by vehicle-mounted apparatus. Spraying and fogging often fail to target indoor resting mosquitoes (6,7). Nevertheless adulticides remain important in combating epidemics and the high visibility of this intervention can sometimes favour its deployment, especially at times when mosquito numbers are particularly high. The pyrethroids are the usual class of insecticide used in space spraying, due to their rapid knockdown action and low mammalian toxicity. However, some dengue control programmes continue to use organophosphates (malathion, pirimiphos methyl or fenitrothion) to target adult mosquitoes. Recent trials using pyrethroid treated material such as curtains or container covers showed promise and may represent a more effective means of adulticiding (8).
The evolution and spread of resistance to insecticides is a major concern for the control of all arthropod transmitted infections and dengue is no exception. Indeed the reliance by most dengue control programmes on just two of the four classes of insecticide available for use in public health, poses additional selection pressure on the mosquito vectors.
The objective of this review is to determine and describe the current distribution of insecticide resistance in dengue vectors and to evaluate the impact of resistance on Aedes control activities. For the purpose of the review we have included reports of resistance to all four major classes of chemical insecticides (carbamates, organochlorines, organophosphates and pyrethroids) but the discussion will focus primarily on those insecticides most widely employed for dengue control. We will describe alternative approaches used to monitor for resistance and call for greater standardization in methodologies and data reporting. Finally, we will identify some important knowledge gaps that need to be addressed by further research.
Methods
We looked for studies of any design, in which any of the named insecticides were used against Ae. aegypti or Ae. albopictus mosquitoes and in which insecticide resistance was a reported outcome.
Inclusion criteria
Public databases (Web of Science and PubMed) were searched with the key words 'Aedes aegypti' or 'Aedes albopictus' and one of the following terms: 'insecticide resistance', 'DDT', 'pyrethroids', 'carbamates', 'organophosphates' or 'organochlorines'. Insecticide resistance in other regional secondary dengue vectors was not included in the search strategy. Manuscripts in all languages were considered. Abstracts were retrieved and, if necessary, translated into English by a native speaker. All abstracts were read and full manuscripts were retrieved for those papers reporting data on the distribution, impact, or causes of insecticide resistance in dengue vectors.
Exclusion criteria
For studies reporting insecticide bioassay data, we excluded results where the mosquitoes had been subjected to artificial selection with insecticide in the laboratory (although data on resistance mechanisms were extracted from these manuscripts). We also rejected any bioassay results where fewer than 20 mosquitoes from each population were bioassayed. Plans to exclude studies where adequate controls were not reported were forsaken, due to the variability in the methodologies employed by the different studies (described in detail below). Publications from all years were considered and included in the database but where bioassay data were available for multiple years for the same study site only the most recent data set was used to generate the maps.
Results
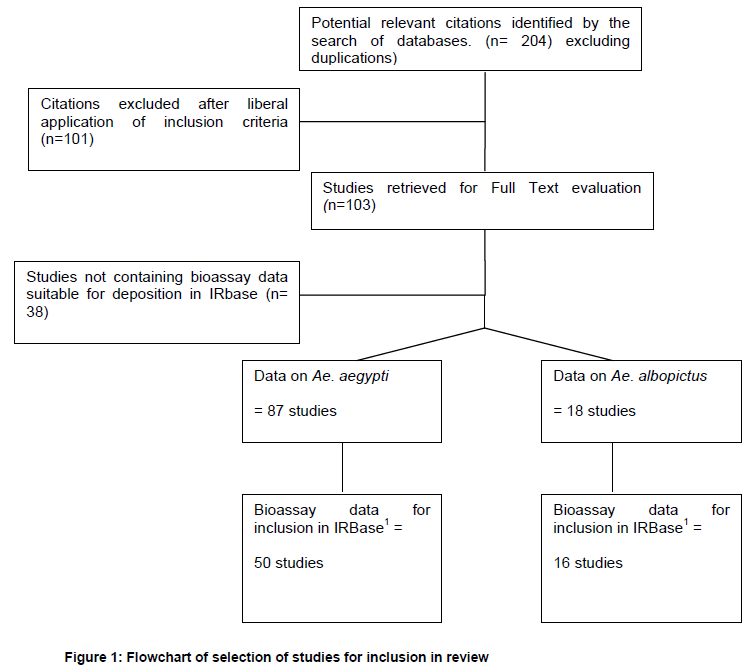
A flowchart showing the number of studies included in this review is given in Figure 1.
A total of 204 manuscripts were retrieved. Nearly half (101) were excluded either because insufficient detail was provided on the methodologies employed, they did not contain any new data (review articles fell into this category) or they did not contain information relevant to the topic of this review. Of the 103 papers included, 87 contained data on Ae. aegypti, 18 on Ae. albopictus and two manuscripts contained data on both species. Forty nine of the 87 manuscripts included for Ae. aegypti and 16 of the 18 included for Ae. albopictus contained bioassay data that was extracted for deposition in IRbase. IRbase is a database for recording information on insecticide resistance in disease vectors. It is accessible via vectorbase.org. The remainder either reported data from laboratory selected strains only or the sample sizes were too low (less than 20 mosquitoes bioassayed). Information on the resistance mechanisms and the impact of resistance on dengue control operations was extracted from studies included in the review but not necessarily containing bioassay data suitable for deposition in IRbase.
Geographical distribution of included studies
At the continental scale, the distribution of published studies on the susceptibility of dengue vectors to insecticides reflects the burden of disease, with the vast majority of reports from Asia and South America and very few from Europe or Africa. However a closer look reveals a clear distortion, with the majority of the reports originating from a handful of countries (Table 1). Amongst these countries are Brazil and Cuba. Both these countries have well-structured national programmes for resistance monitoring. However, data from other countries with similar national resistance monitoring activities, such as Vietnam and Colombia, are not readily accessible via searches of global literature databases as these data are currently retained in country without being released into the public domain.
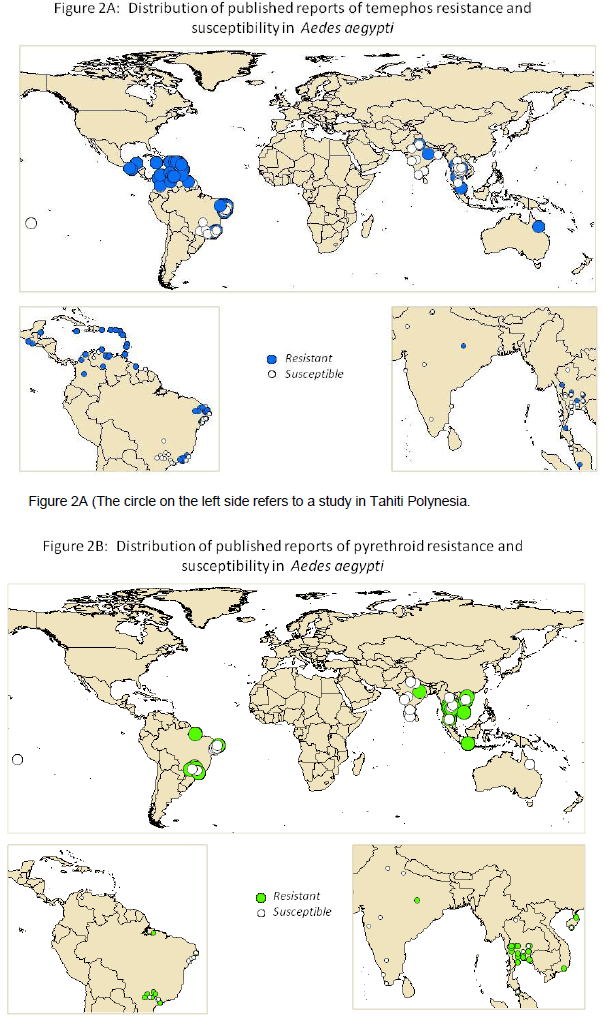
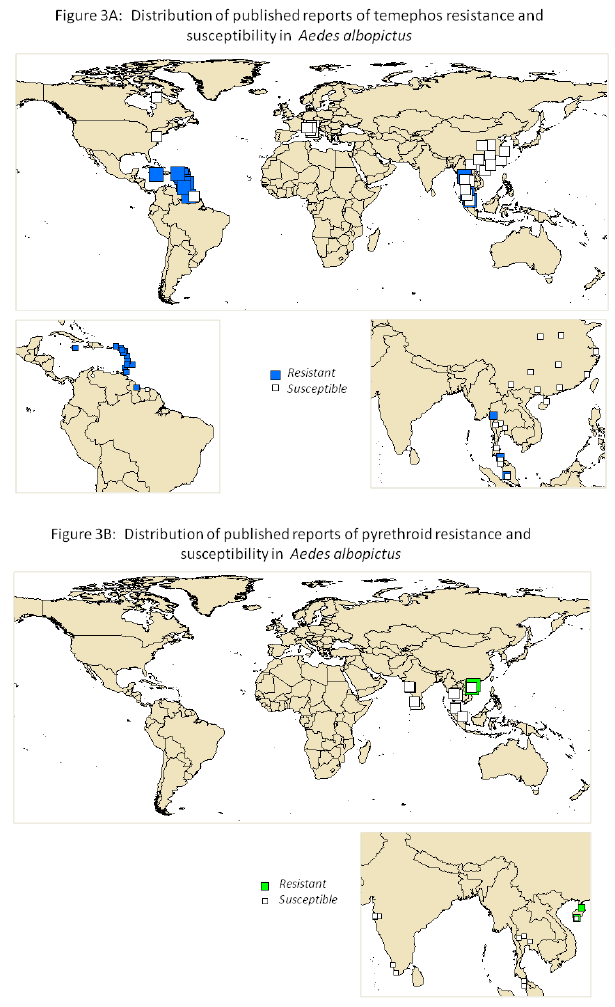
Some studies consider resistance to a single insecticide in either the larval or adult stages. Others provide comprehensive data on the susceptibility of the local vector population to a range of different insecticides. In order to capture all the available information the bioassay data for both life stages and all four classes of insecticide were extracted from 65 published studies in a format suitable for uploading into a public database. This data is included as supplementary information. The information on the distribution of resistance to the insecticides most widely used for control of dengue vectors today, is summarized in Figures 2 and 3.
As described below, there was wide variation in the methodologies used to record and interpret susceptibility data making cross comparisons between studies very difficult. Hence no attempt has been made to indicate the level of resistance in the local vector populations in the figures. For study sites with more than one year of data only the most recent dataset was used to derive the maps.
Figures 2 and 3 reveal that resistance to pyrethroids was detected in the majority of published studies on this topic and that temephos resistance is also widespread. However, given that there is likely to be a clear publication bias towards 'positive' reports of resistance, this should be interpreted with caution. Nevertheless it is apparent that insecticide resistance is widespread in Ae.aegypti and also present in some populations of Ae. albopictus, although there are very few published reports of bioassays on the latter species. Another striking finding is that insecticide resistance is often very focal. For example the percentage mortality of Ae. aegypti females after exposure to a diagnostic dose of malathion varied from 43 to 100% in different counties in Sao Paolo State in Brazil (9). In Nakhon Sawan, North central Thailand, resistance to temephos varied from 50 to 100% mortality in local Ae. aegypti populations sampled throughout the province (10). Furthermore, work in the state of Quintana Roo, Mexico illustrates that even the mechanisms of resistance can be very focal (11).
Variations in methodologies used for resistance monitoring
There are two commonly used insecticide bioassay methodologies. In the first, insects are exposed to a range of insecticide concentrations and the amount of insecticide needed to kill 50 or 95% of the population (LC50 or LC95) is reported. Usually a known susceptible strain is run in parallel and the values for the field populations are divided by those for the standard susceptible strain to obtain a resistance ratio (RR). The second, simpler, approach uses a single diagnostic dose of insecticide that has been previously shown to kill 100% of a susceptible population. Deviations from 100% mortality are taken as indications of resistance, with a cut off of less than 80% mortality usually being employed to describe a resistant population. Both of these methods were utilized by studies in this review but modifications of the original protocols, or variations in the way in which the results were interpreted, greatly complicated cross study comparisons. Some of the sources of discrepancy are listed below:
Changes in the World Health Organization guidelines not applied in all the studies. Two separate documents giving diagnostic dosages of insecticides for a range of vector species have been produced by WHO (12,13). The diagnostic doses for Ae. aegypti for several insecticides were reduced in the revised 1992 guidelines (for example, from 0.02 mg/l temephos in 1981 report to 0.012 mg/l in 1992). Despite this revision, several studies are still using the original, higher doses to determine the susceptibility of their local vectors (e.g. 14,15).
Absence of guidelines on diagnostic doses for some insecticides. Diagnostic doses have not been included for many of the insecticides commonly used in dengue control programmes in the WHO guidelines and this has lead to large discrepancies in the methodologies. For example, Sharma et al (15) exposed adult Ae. aegypti from India to 1% fenitrothion for two hours, whereas in Thailand, Jirakanjanakit et al (16) used a one-hour exposure to 0.5% fenitrothion. Both studies present results as percentage mortality but data between the studies cannot be easily compared.
Independent determination of diagnostic dose. In some cases, diagnostic doses were first calculated by measuring the dose response of a susceptible strain. This was used to establish a single diagnostic dose that is typically double the concentration needed to kill 99% of the susceptible strain (as recommended by WHO (17)). This dose is then used to assay the response of field populations. For example, a study in Thailand used 0.04mg/l temephos as a diagnostic dose for Ae. aegypti (10). As this dose is over 3-fold higher than the WHO diagnostic dose, these mortality results are presumably much higher than would have been obtained with a more standardized study design.
Wide variation in the LC50s of the susceptible strain affect calculation of resistance ratios. The RR of a population is clearly dependent on the LC50 or LC95 value obtained for the susceptible strain. Wide variations in these values were noted between the studies, even when the same laboratory susceptible strain was used. For example, the LC50 values for temephos for the Rock strain varied >5-fold between different studies (18, 19). This is less of an issue when actual LC50 values are presented but if data are summarized as RR these variations will confound interpretation of the results.
Causes of insecticide resistance
Bioassays can only detect resistance when it is already established in a population. Tools to detect resistance when it first appears would enable timely resistance management actions to be taken which, if successful, would prevent resistance compromising control activities. Development of diagnostic tools for insecticide resistance requires knowledge of resistance mechanisms. Alterations in the molecular target sites of insecticides, which reduce the binding of insecticides are the most
understood resistance mechanisms. Several mutations in the sodium channel, the target site of DDT and pyrethroid insecticides, have been reported in Ae. aegypti (20). Two alternative substitutions at one of the polymorphic sites, residue 1016, have been linked to pyrethroid resistance and recently, methodologies to detect these mutations (often referred to as kdr mutations) in individual mosquitoes have been reported (21, 22). These kdr assays are already being employed to monitor the effect of control strategies on insecticide resistance in several countries in Latin America. However, until these molecular assays for target site resistance are complemented by similar assays to detect other resistance mechanisms (most importantly metabolic resistance), molecular tools cannot be used as a substitute for bioassays. The sequencing of the Ae. aegypti genome (23) has advanced research on insecticide resistance mechanisms and it is hoped that ongoing research will be fruitful in the search for additional diagnostic markers of insecticide resistance in dengue vectors.
Biochemical assays to detect alterations in activities of enzyme families associated with insecticide metabolism, have been available for over two decades and are being routinely used in resistance monitoring programmes in Brazil, Cuba and Colombia amongst others. These assays can provide important mechanistic data for predicting patterns of cross resistance and thereby suitable alternative chemicals. However, the lack of sensitivity and specificity of some of the assays, and difficulties in interpreting the data, are major obstacles to their routine implementation. Similarly, synergists such as piperonyl butoxide and diethyl maleate, which block the activity of enzymes responsible for insecticide detoxification, can be used to explore the role of metabolic resistance (24, 25). If resistance is due to elevated insecticide metabolism addition of an appropriate synergist should reduce the LC50 to the level of the susceptible strain. Often, the application of synergists to study insecticide resistance mechanisms is limited by the need for large numbers of live insects. Furthermore, as with the biochemical assays, sensitivity and specificity issues necessitate the use of caution when interpreting the use of synergist data.
Impact of current levels of insecticide resistance on vector control
Only a small number of studies address the impact of insecticide resistance on mosquito control in the field. A comparison of Caribbean populations of Ae. aegypti found that the effectiveness of temephos applied to water storage containers at field doses was compromised by insecticide resistance. Although the susceptible reference population continued to suffer 100% mortality over the eight weeks of the trial, control of the three field resistant populations was compromised by week three (26). Similarly in Brazil, water containers treated with temephos at the field dose and left to weather outside achieved less than 70% mortality of local resistant populations of Ae. aegypti after just four weeks whereas complete control of the susceptible strain was achieved throughout the seven week duration of the trial (27). Other studies (28) also tested the response of local mosquito populations against the field application dose of temephos (1mg/l) but, rather than measuring the residual effect of the insecticide treatment, they reported cumulative mortality over a two-hour period and hence provide little information about the efficacy of local control measures.
Several studies have demonstrated successive increases in resistance to pyrethroid insecticides over time in dengue vectors, often following repeated applications of this insecticide class for dengue control (9,29,30). Furthermore, a cage-bioassay in the Caribbean island of La Martinique demonstrated that the efficacy of deltamethrin applied by a vehicle thermal fogger was compromised by resistance in the local Ae. aegypti population (31).
Discussion
In the process of undertaking this review, it became apparent that there is a very large amount of data on insecticide resistance in dengue vectors that is not captured in the published literature. Several national control programmes retain large volumes of data on resistance levels in local vectors that are not publically accessible. This poses a problem for the selection of appropriate insecticides for dengue control. Aedes aegypti and to an increasing extent Ae. albopictus, are cosmotropical species which do not respect country borders. Hence it is important for dengue control programmes to be aware of any reports of resistance hindering control efforts within the region. A centralized database for recording insecticide resistance in a standardized format would greatly facilitate exchange of information and future planning of insecticide based interventions. Fortunately a database (IRbase) has now been developed specifically for this purpose and is freely accessible on the internet (via vectorbase.org) (32). The data extracted from published literature and used in the writing of this review have already been deposited in this database and the challenge now is to encourage universal utilization of this resource. Furthermore, in order to maximize the utility of IRBase, greater attention needs to be paid to the way in which insecticide resistance data are recorded. For example it is important that, when using a range of insecticide concentrations or exposure times, the LC50/LT50 and LC90/LT90 values are calculated for a laboratory susceptible strain as well as for the population under study and that the actual values are reported rather than being summarized as resistance ratios. In addition, diagnostic doses still need to be established for some insecticides that are used in dengue control and all entomologists should monitor the WHO pesticide evaluation scheme (WHOPES) website (http://www.who.int/whopes/guidelines/en/) for alterations or additions to the guidelines on diagnostic doses.
Even considering the limitations in the available data, it is evident that there are still large areas of the world where insecticides are being used for dengue control in the absence of any data on the susceptibility of local vector populations. Given that countries with effective resistance monitoring programmes are reporting alarming increases in the level of insecticide resistance, this information deficit should be urgently addressed.
A small number of studies have demonstrated a reduced efficacy of mosquito control as a result of insecticide resistance and in some cases has led to a change in control policy (27), but further studies are needed on the impact of pyrethroid resistance on the efficacy of adulticides.
Furthermore, there have been no studies to examine the impact of insecticide resistance on dengue transmission. Hence, although insecticide resistance is often stated as an impediment for dengue control, there appears to be little evidence to substantiate this. This is no cause for complacency. Instead it suggests that the correct studies have not been designed and implemented to determine the operational impact of insecticide resistance. However, clearly in order to persuade control programmes of the necessity of regular insecticide resistance monitoring, further work is needed to demonstrate the impact of resistance on current control activities and to demonstrate that resistance management strategies can be effective.
Resistance management strategies generally recommend the rotation of chemicals with different modes of action and the use of non-chemical methods of control. The implicit assumption is that resistance to a chemical will disappear from a population once the selection pressure is removed. However, for this to be successful the selection pressure must be removed before resistance becomes established in the population (indeed a truly effective resistance management strategy would rotate among insecticides at sufficient frequency to avoid the emergence of resistance at all). Unfortunately, dengue control programmes that have changed insecticide classes in response to resistance have not always witnessed a decrease in resistance to the original insecticide. For example in Singapore, high levels of resistance to permethrin still persist in dengue vectors nine years after its replacement with pirimiphos methyl as the adulticide for dengue control (33). This may partly be attributed to domestic use of pyrethroid insecticides by households, but it should also be noted that metabolic resistance mechanisms can cause cross resistance across insecticide classes ( 34). Similarly, in Brazil, the interruption of temephos use did not result in a rapid return to susceptibility (27).
The poor results from resistance management interventions highlight the need for more effective tools to detect resistance when it first appears in a population, and for new chemicals to use as substitutes when resistance impacts on control activities. Both of these requirements are being addressed by the Innovative Vector Control Consortium, a public-private partnership which is developing new tools for control of the mosquito vectors (35) but it is recognized that it will be at least a decade before new active ingredients are available for mosquito control.
Finally, very little is known about the relative contribution of global migration versus de novo mutations in shaping the current distribution of insecticide resistance in dengue vectors or of the role that dengue control activities are playing in selecting for resistance. These are not just academic questions. The chances of implementing effective policy changes to mitigate resistance can be greatly improved if resistance is a local occurrence arising from the choice of insecticide used in dengue control operations. However, if external factors, such as the use of domestic insecticidal products (e.g. aerosols) and migration of resistant mosquitoes from neighboring populations or from agricultural areas, are shaping the profile of resistance then the ability to manage resistance is reduced.
Supplementary tables
Table S1. Data extracted from published literature on insecticide resistance in Ae aegypti. Insecticide resistance data have been recorded in a format compatible with submission to IRbase (http://anobase.vectorbase.org/ir/)
Table S2: Data extracted from published literature on insecticide resistance in Ae albopictus. Insecticide resistance data have been recorded in a format compatible with submission to IRbase (http://anobase.vectorbase.org/ir/)
Acknowledgements
The authors of this review are deeply indebted to Dr Louise Kelly-Hope for her invaluable work in producing the maps that document the distribution of resistance.
Summary boxes
What has been learned from this review?
- Resistance to all four classes of insecticide (carbamates, organochlorines, organophosphates and pyrethroids) has developed in Aedes aegypti
- Temephos resistance is compromising the success of larviciding in some countries and there is evidence that pyrethroid resistance is negatively impacting on adulticide campaigns
- Insecticide resistance can be localized to specific areas
Implications for public health practice
- Resistance monitoring must be routinely undertaken in all dengue control programmes utilizing insecticides
- Careful attention should be paid to how resistance is reported to ensure that comparative analysis can be conducted between different areas and time points
- Resistance management strategies need to be implemented to avoid compromising control.
- Non-chemical means of Aedes control (e.g. removal of breeding sites, use of predators) should be employed wherever possible.
Priority research areas
- Standardization of protocols and reporting procedures for insecticide resistance in dengue vectors
- Improved means of disseminating and accessing resistance data
- New molecular tools to detect insecticide resistance when it first appears in a population
- Studies into the factors influencing the evolution and distribution of insecticide resistance
- Further studies on the relationship between insecticide resistance and efficacy of insecticide based interventions used to control dengue epidemics.
References
1. World Health Organization (2008). Dengue and dengue haemorrhagic fever. Fact sheet 117 [ Links ]
2. Kyle JL, Harris E (2008). Global spread and persistence of dengue. Annu Rev Microbiol; 62:71-92. [ Links ]
3. McCall PJ, Kittayapong, P (2006). Control of dengue vectors: tools and strategies. Scientific Working Group, World Health Organization [ Links ]
4. World Health Organization (2006). Pesticides and their application for the control of vectors and pests of public health importance. 6th ed. Geneva, WHO/CDS/NTD/WHOPES.GCDPP/2006.a [ Links ]
5. World Health Organization (2007). Expert consultations for the 4th edition of the guidelines for drinking water safety. WHO/HSE/WSH/07/11 [ Links ]
6. Castle T, Amador M, Rawlins S, Figueroa JP, Reiter P (1999). Absence of impact of aerial malathion treatment on Aedes aegypti during a dengue outbreak in Kingston, Jamaica. Rev Panam Salud Publica; 5:100-5. [ Links ]
7. Perich MJ, Davila G, Turner A, Garcia A, Nelson M (2000). Behavior of resting Aedes aegypti (Culicidae : Diptera) and its relation to ultra-low volume adulticide efficacy in Panama City, Panama. J Med Entomol; 37(4): 541-546. [ Links ]
8. Kroeger A, Lenhart A, Ochoa M, Villegas E, Levy M, Alexander N, McCall PJ (2006). Effective control of dengue vectors with curtains and water container covers treated with insecticide in Mexico and Venezuela: cluster randomised trials. BMJ; 332(7552):1247-1252. [ Links ]
9. Macoris MDD, Andrighetti MTM, Otrera VCG, de Carvalho LR, Caldas AL, Brogdon WG (2007). Association of insecticide use and alteration on Aedes aegypti susceptibility status. Mem Inst Oswaldo Cruz;102(8):895-900. [ Links ]
10. Jirakanjanakit N, Saengtharatip S, Rongnoparut P, Duchon S, Bellec C, Yoksan S (2007). Trend of Temephos resistance in Aedes (Stegomyia) mosquitoes in Thailand during 2003-2005. Environ Entomol; 36(3):506-511. [ Links ]
11. Flores AE, Grajales JS, Salas IF, Garica GP, Becerra MHL, Lozano S, Brogdon WG, Black WC, Beaty B (2006). Mechanisms of insecticide resistance in field populations of Aedes aegypti (L.) from Quintana Roo, Southern Mexico. J Am Mosq Control Assoc; 22(4):672-677. [ Links ]
12. World Health Organization (1981). Instructions for determining the susceptibility or resistance of mosquito larvae to insecticides. WHO/VBC/81.807 [ Links ]
13. World Health Organization (1992). Vector resistance to pesticides. WHO/TRS/818 [ Links ]
14. Romi R, Toma L, Severini F, Di Luca M (2003). Susceptibility of Italian populations of Aedes albopictus to temephos and to other insecticides. J Am Mosq Control Assoc;19(4):419-423. [ Links ]
15. Sharma S N, Saxena VK, Lal S (2004). Study on susceptibility status in aquatic and adult stages of Aedes aegypti and Ae. albopictus against insecticides at international airports of South India. J Commun Dis;36(3):177-181. [ Links ]
16. Jirakanjanakit N, Rongnoparut P, Saengtharatip S, Chareonviriyaphap T, Duchn S, Bellec C, Yoksan S (2007). Insecticide susceptible/resistance status in aedes (Stegomyia) aegypti and Aedes (Stegomyia) albopictus (Diptera : Culicidae) in Thailand during 2003-2005. J Econ Entomol;100(2): 545-550. [ Links ]
17. World Health Organization (2005). Guidelines for laboratory and field testing of mosquito larvicides. WHO/CDS/WHOPES/GCDPP/2005.13 [ Links ]
18. Rodriguez MM, Bisset JA, Fernandez D (2007). Levels of insecticide resistance and resistance mechanisms in Aedes aegypti from some Latin American countries. J Am Mosq Control Assoc;23(4):420-429. [ Links ]
19. Ponlawat A, Scott JG, Harrington LC (2005). Insecticide susceptibility of Aedes aegypti and Aedes albopictus across Thailand. J Med Entomol;42(5):821-825. [ Links ]
20. Brengues C, Hawkes NJ, Chandre F, McCarroll L, Duchon S, Guillet P, Manguin S, Morgan JC, Hemingway J (2003). Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med Vet Entomol;17(1):87-94. [ Links ]
21. Saavedra-Rodriguez K, Urdaneta-Marquez L, Rajatileka S, Moulton M, Flores AE, Fernandez-Salas I, Bisset J, Rodriguez M, McCall PJ, Donnelly MJ, Ranson H, Hemingway J, Black WC (2007). A mutation in the voltage-gated sodium channel gene associated with pyrethroid resistance in Latin American Aedes aegypti. Insect Mol Biol; 16(6): 785-798. [ Links ]
22. Rajatileka S, Black IV WC, Saavedra-Rodriguez K, Trongtokit Y, Apiwathnasorn C, McCall PJ, Ranson H (2008). Development and application of a simple colorimetric assay reveals widespread distribution of sodium channel mutations in Thai populations of Aedes aegypti. Acta Trop; 108(1):54-57. [ Links ]
23. Nene V , Wortman JR , Lawson D , Haas B , Kodira C , Tu ZJ , Loftus B , Xi Z , Megy K , Grabherr M , Ren Q , Zdobnov EM , Lobo NF , Campbell KS , Brown SE , Bonaldo MF , Zhu J , Sinkins SP , Hogenkamp DG , Amedeo P , Arensburger P , Atkinson PW , Bidwell S , Biedler J , Birney E , Bruggner RV , Costas J , Coy MR , Crabtree J , Crawford M , Debruyn B , Decaprio D , Eiglmeier K , Eisenstadt E , El-Dorry H , Gelbart WM , Gomes SL , Hammond M , Hannick LI , Hogan JR , Holmes MH , Jaffe D , Johnston JS , Kennedy RC , Koo H , Kravitz S , Kriventseva EV , Kulp D , Labutti K , Lee E , Li S , Lovin DD , Mao C , Mauceli E , Menck CF , Miller JR , Montgomery P , Mori A , Nascimento AL , Naveira HF , Nusbaum C , O'leary S , Orvis J , Pertea M , Quesneville H , Reidenbach KR , Rogers YH , Roth CW , Schneider JR , Schatz M , Shumway M , Stanke M , Stinson EO , Tubio JM , Vanzee JP , Verjovski-Almeida S , Werner D , White O , Wyder S , Zeng Q , Zhao Q , Zhao Y , Hill CA , Raikhel AS , Soares MB , Knudson DL , Lee NH , Galagan J , Salzberg SL , Paulsen IT , Dimopoulos G , Collins FH , Birren B , Fraser-Liggett CM , Severson DW (2007). Genome sequence of Aedes aegypti, a major arbovirus vector. Science; 316(5832):1718-1723. [ Links ]
24. Young SJ, Guning RV, Moores GD (2005). The effect of piperonyl butoxide on pyrethroid-resistance-associated esterases in Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Pest Manag Sci;61: 397- 401. [ Links ]
25. Pasay C , Arlian L , Morgan M , Gunning R , Rossiter L , Holt D , Walton S , Beckham S , McCarthy J (2009). The effect of insecticide synergists on the response of scabies mites to pyrethroid acaricides. PLoS Negl Trop Dis;3(1): e354. [ Links ]
26. Rawlins SC (1998). Spatial distribution of insecticide resistance in Caribbean populations of Aedes aegypti and its significance. Rev Panam Salud Publica;4(4): 243-251. [ Links ]
27. Montella IR, Martins AJ, Viana-Medeiros PF, Lima JBP, Braga IA, Valle D (2007). Insecticide resistance mechanisms of Brazilian Aedes aegypti Populations from 2001 to 2004. Am J Trop Med Hyg; 77(3):467-477. [ Links ]
28. Chen CD, Nazni WA, Lee HL, Sofian-Azirun M (2005). Susceptibility of Aedes aegypti and Aedes albopictus to temephos in four study sites in Kuala Lumpur City Center and Selangor State, Malaysia. Trop Biomed; 22(2):207-216. [ Links ]
29. da-Cunha MP, Lima JBP, Brogdon WG, Moya GE, Valle D (2005). Monitoring of resistance to the pyrethroid cypermethrin in Brazilian Aedes aegypti (Diptera : Culicidae) populations collected between 2001 and 2003. Mem Inst Oswaldo Cruz;100(4):441-444. [ Links ]
30. Mekuria Y, Gwinn TA, Williams DC, Tidwell MA (1991). Insecticide susceptibility of Aedes aegypti from Santo Domingo, Dominican Republic. J Am Mosq Control Assoc;7(1):69-72. [ Links ]
31. Marcombe S, Carron A, Darriet F, Etienne M, Agnew P, Tolosa M, Yp-Tcha MM, Lagneau C, Yebakima A, Corbel V(2009). Reduced efficacy of pyrethroid space sprays for dengue control in an area of Martinique with pyrethroid resistance. Am J Trop Med Hyg;80(5):745-751. [ Links ]
32. Dialynas E, Topalis P, Vontas J, Louis C (2009). MIRO and IRbase: IT tools for the epidemiological monitoring of insecticide resistance in mosquito disease vectors.PLoS Negl Trop Dis;3(6):e465. [ Links ]
33. Ping LT, Yatiman R, Gek LPS (2001). Susceptibility of adult field strains of Aedes aegypti and Aedes albopictus in Singapore to pirimiphos-methyl and permethrin. J Am Mosq Control Assoc; 17(2):144-146. [ Links ]
34. Rodríguez MM, Bisset JA, Ruiz M, Soca A (2002). Cross-resistance to pyrethroid and organophosphorus insecticides induced by selection with temephos in Aedes aegypti (Diptera: Culicidae) from cuba. J Med Entomol;39(6):882-888. [ Links ]
35. Hemingway J, Beaty BJ, Rowland M, Scott TW, Sharp BL (2006). The innovative vector control consortium: improved control of mosquito-borne diseases. Trends Parasitol; 22(7):308-312. [ Links ]
Submitted: November 2008; Accepted: June 2009; Published: December 2009
* Corresponding author: Hilary Ranson, hranson@liverpool.ac.uk
{kind=link}
{kind=link}
{kind=link}
{kind=link}